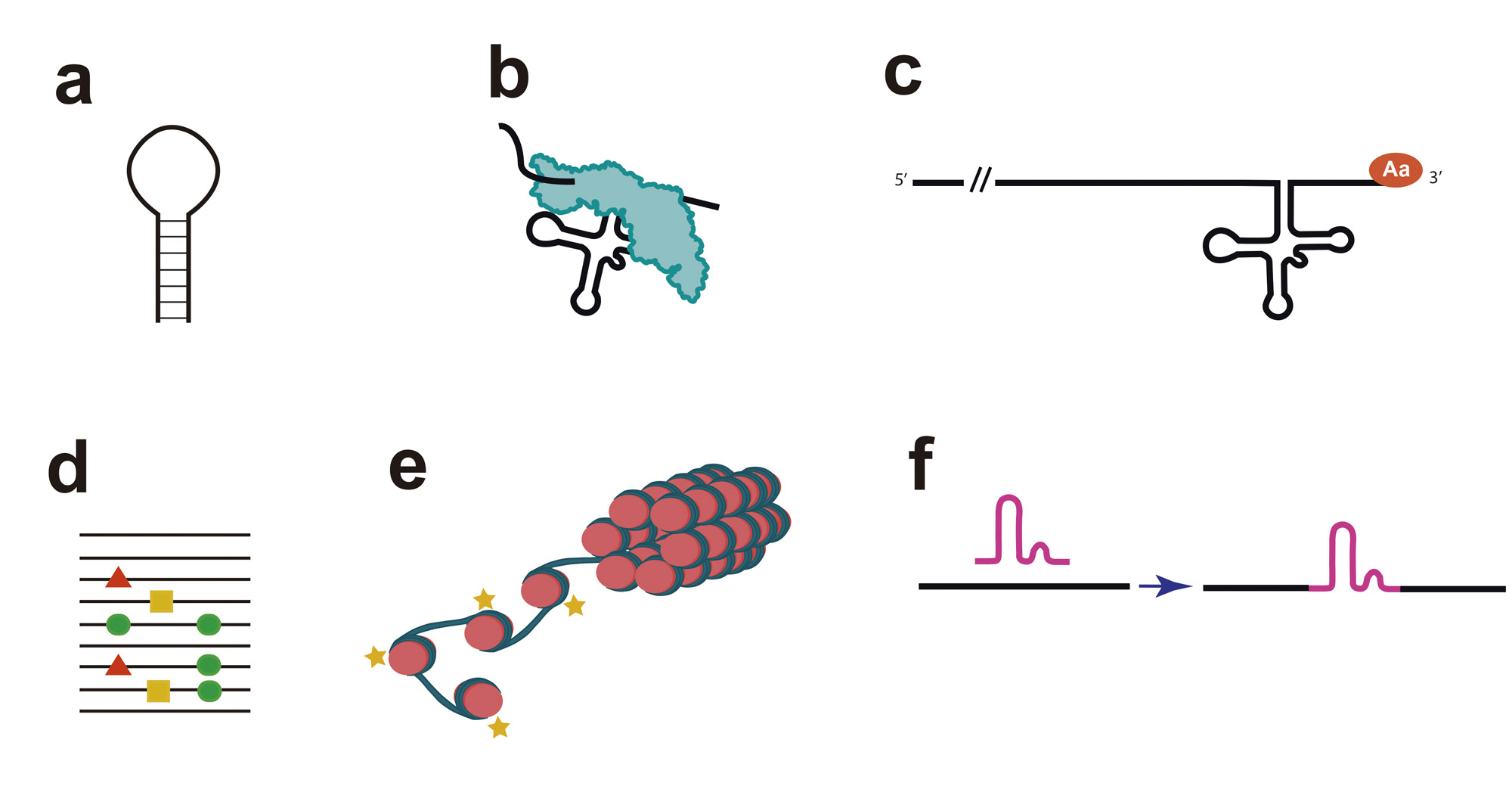Arqueología del RNA codificante
Ascensión Ariza-Mateos,1,2Carlos Briones,3,4Celia Perales,2,4,5Esteban Domingo,2,4and Jordi Gómez1,4
1 Laboratory of RNA Archaeology, Instituto de Parasitología y Biomedicina "López-Neyra" (CSIC), Granada, Spain. 2 Centro deBiología Molecular "Severo Ochoa" (CSIC-UAM), Campus de Cantoblanco, Madrid, Spain. 3 Department of Molecular Evolution, Centro de Astrobiología (CSICINTA), Madrid, Spain. 4 Centro de Investigación Biomédica en Red deEnfermedades Hepáticas y Digestivas (CIBERehd), Instituto de Salud Carlos III, Madrid, Spain. 5 Department of Clinical Microbiology, IIS-Fundación Jiménez Díaz, UAM, Madrid, Spain.
Enlace a la nota de prensa elaborada por el CSIC ... 
RESUMEN
A partir de los resultados experimentales y modelos realizados en nuestros laboratorios (IPBLN, CAB y CBMSO), hemos desarrollado una hipótesis para aproximarnos al pasado del RNA durante las primeras fases de la evolución bioquímica, que hemos denominado “arqueología del RNA”. Se trata de un método, alternativo y complementario a la filogenia molecular, para buscar elementos de RNA del pasado, con independencia de que sus secuencias se hayan difuminado y su función haya cambiado. Estos elementos son aquellos que han sido reprimidos, desdibujados o re-organizados, y por tanto representan a “los perdedores” en los conflictos que han determinado la complexificación de la vida. Es interesante destacar que dichos elementos de RNA han sido tradicionalmente olvidados en los estudios de la historia evolutiva de esta macromolécula genética y funcional.
Los factores empleados en el estudio arqueológico, nuestra "caja de herramientas”, han sido la ribozima RNasa P humana y bacteriana, y la enzima RNAsa III de E. coli. Las primeras procesan los precursores del tRNA y la segunda los RNAs largos (≥20 nucleótidos), pero ambas actividades pueden identificar además moléculas estructuralmente similares al tRNA o al RNA de doble cadena. También se ha empelado un tercer factor, la radiación ultravioleta C (UV-C), que es capaz de activar una auto-escisión oxidativa específica en algunas estructuras locales terciarias del RNA. Todos estos factores remiten de una u otra forma al pasado remoto de la vida. El “terreno” estudiado ha sido el conjunto de RNAs mensajeros hepáticos, el mRNA del virus de la hepatitis C (VHC) y otros virus evolutivamente relacionados como miembros de las familias Flaviviridae y Picornaviridae.
Con los ensayos realizados se analiza la capacidad del RNA para reconocer o ser reconocido por estos factores, lo que nos informa directamente sobre la materialidad de los RNAs sustratos en cuestión. Esto se diferencia de los métodos filogenéticos, que basan sus análisis en representaciones ya que comparan las secuencias de bases del RNA (símbolos) o, los mapas y planos de sus estructuras.
Las RNasas P detectan numerosos elementos en el interior de los mRNAs celulares, especialmente en los ligados al metabolismo del RNA y al metabolismo energético, que representan los tipos de RNAs codificantes más primitivos de la célula. También identifican elementos susceptibles en el mRNA del VHC y otros virus relacionados. En los ejemplos que hemos estudiado, la actividad RNasa III reconoce elementos situados en las proximidades de los elementos identificados por la RNasa P, y estos “conjuntos de elementos” se encuentran actualmente en zonas coincidentes con regiones funcionales de los mRNA virales o celulares. Esto nos ha llevado, por un lado, a sustentar modelos teóricos en los que los elementos parecidos al tRNA concatenados podrían estar en el origen de los mRNAs. Y también nos permite apoyar la hipótesis de que el RNA de origen bacteriano 10S podría ser un “bloque de construcción” original de los mRNAs al combinar también elementos dsRNA/tRNA.
Además de los ensayos discutidos, otros puntos de partida en el análisis de las interacciones y dinámicas moleculares protagonizadas por el RNA, como las que se nombran en la Figura se acomodan bien a esta línea de investigación arqueológica que se inicia con este trabajo.
Enlace al artículo: https://nyaspubs.onlinelibrary.wiley.com/doi/10.1111/nyas.14173
Pie de Figura. A) Los tallos-lazo de los RNAs son la forma básica del plegamiento del RNA y permite la asociación con otros para formar estructuras complejas cuando la interacción es a nivel intramolecular, o bien la formación de consorcios de RNA cuando las interacciones son intermoleculares. B) La ribozima RNasa P (azul) procesa el precursor del RNA. C) Genomas de los virus de RNA que infectan plantas contienen un elemento parecido al tRNA que es capaz de cargarse específicamente con un Aa. (D) representación esquemática de la complejidad de una cuasiespecies virales. Las secuencias virales se representan como líneas, y las mutaciones puntuales como símbolos coloreados. E) representación del cambio entre cromatina activa o inactiva a través de las marcas epigenéticas. F) elementos autoreplicativos que invaden el genoma del huésped.
|