|
LINEAS DE INVESTIGACIÓN
FUNDAMENTOS CIENTÍFICOS - DNA MÓVIL EN HUMANOS
El genoma humano ha sido invadido por varias olas de elementos móviles de DNA que se han multiplicado y posteriormente decaído a lo largo de la evolución, dejando incontables copias fósiles como consecuencia de su pasado replicativo. Estas copias representan alrededor de la mitad del genoma y se encuentran ya consolidadas como elementos estructurales, reguladores e, incluso, ocasionalmente codificantes (Lander et al. Nature 2001). Dentro de este grupo polifilético de elementos, los retrotransposones son los que están/han estado activos en tiempos recientes. Estos elementos se transcriben a un RNA intermediario que es reversotranscrito de nuevo a una nueva copia de DNA (Kazazian. FT Press Science 2011). En este grupo, los retrovirus endógenos (ERV; estrechamente relacionados con retrovirus) estuvieron activos hasta muy recientemente (y aún lo están en otros organismos como Drosophila, ratón y cerdo) (Belshaw et al. J Virol 2005; Maksakova et al. PLoS Genet 2006). Solo los elementos LINE-1 (L1), Alu y SVA (todos no-ERV), son todavía móviles en humanos, causando mutagénesis insercional tanto en la línea germinal como en células somáticas, aunque de forma limitada gracias al control epigenético. Estos elementos crean diversidad genética a razón de ~1 inserción heredable por cada 40 nacimientos, causando ocasionalmente casos de enfermedades genéticas quizá a una escala evolutivamente tolerable (Ewing and Kazazian. Genome Res 2010; Feusier et al. Genome Res 2019; Kazazian and Moran. N Engl J Med 2017). Sin embargo, otros autores y nosotros hemos mostrado que pueden desregularse en cáncer y su influencia en la diversidad genética del mismo puede marcar su progresión y metástasis (Scott et al. Genome Res 2016; Tubio et al. Science 2014; Nguyen et al. Cell Rep 2018).
Un L1 funcional consiste en una secuencia de ~6kb que codifica actividades esenciales para su movilización, incluyendo AP endonucleasa (EN) y reverso transcriptasa (RT). El mecanismo de movilización alterna soportes de DNA y RNA, y está probablemente entroncado en los orígenes evolutivos del actual flujo de información entre ácidos nucleicos (Richardson et al. Mob DNA III 2015). La actividad promotora Pol_II de la 5?UTR es responsable de la transcripción sense de un RNA poliadenilado de L1, que es traducido en el citoplasma y se une en cis a su propia maquinaria. Ambos retornan al núcleo para catalizar la síntesis de una copia nueva por target-primed reverse transcription (TPRT), que implica un corte en una hebra del sitio de inserción del genoma y el uso del extremo 3? hidroxilo como primer para la reversotranscripción del RNA de L1 a la primera hebra del cDNA (Luan et al. Cell 1993; Cost et al. EMBO J 2002). Se desconoce como L1 restaura la integridad de la hebra del genoma y sintetiza la hebra de cDNA complementaria, pero posiblemente implica la coordinación de parte de la maquinaria celular. Por el contrario, Alu y SVA son elementos compuestos, más modernos, pequeños y no codificantes que se movilizan utilizando la maquinaria de L1 en trans (Dewannieux et al. Nat Genet 2003; Hancks et al. Hum Mol Genet 2011).
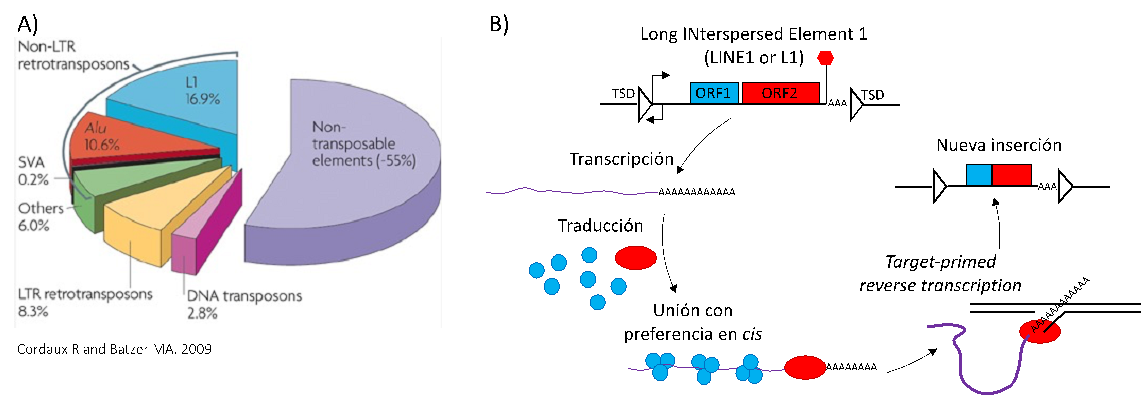
Figure 1. Abundancia de elementos genéticos móviles en humanos y su mecanismo de movilización. A) Gráfico con la abundancia relativa de las principales familias de elementos genéticos móviles o elementos transponibles (Cordaux and Batzter 2009). B) Mecanismo de movilización de elementos L1, que consiste en la transcripción de un L1 completo en un mRNA que es traducido en el citoplasma; que une su maquinaria de transposición en cis; y retorna al núcleo, donde la endonucleasa/transcriptasa reversa ORF2p cataliza la síntesis de una nueva copia mediante transcripción reversa cebada por el DNA genómico del lugar de inserción.
CONTROL EPIGENÉTICO DE DNA MÓVIL Y FORÁNEO EN EL GENOMA HUMANO
El ser humano y el L1 han coevolucionado para alcanzar un nivel de movilización compatible con su común supervivencia. De hecho, se conocen varios eventos en los que los elementos genéticos móviles han influenciado positivamente sus hospedadores, incluyendo humanos (Kapitonov et al. PLoS Biol 2005). El mecanismo central de control de la actividad de los L1HS, los más modernos y los únicos competentes para movilización en humanos, es el silenciamiento epigenético de su promotor, que está mediado por la metilación de una isla CpG presente en su 5?UTR (Muotri et al. Nature 2010). Sin embargo, en el desarrollo embrionario temprano y en el primordio de la línea germinal, ocurren sendas olas de reprogramación que implican una demetilación del DNA generalizada (Greenberg and Bourc?his. Nature 2019). Esto es aparentemente necesario para que las células embrionarias alcancen la pluripotencia en los estadíos tempranos del desarrollo, al tiempo que posibilita una ventana de activación del L1 que le permite alcanzar el genoma de la línea germinal y perpetuar nuevas copias. Es de destacar que existen algo más de 330 copias completas de L1HS en cada individuo, cada una con un promotor con potencial transcripcional. Aproximadamente el 10% de ellos son polimórficos, aún no fijados en la población humana, que añaden a la diversidad genética entre individuos (Ewing and Kazazian. Genome Res. 2011). La mayor parte de estos L1HS quedan rápidamente silenciados por metilación de novo del DNA ya en el embrión pre-implantación, aunque se desconoce el mecanismo con el que las células los localizan y silencian durante la reprogramación.
Las copias de L1HS se han estudiado típicamente en conjunto debido a su extremadamente alta similitud. Su origen replicativo y reciente ha impedido tradicionalmente su estudio individual con técnicas ómicas basadas en lecturas de secuenciación cortas. Durante mi postdoctoral en el laboratorio del Prof. Faulkner (Mater Research Institute, Brisbane, Australia), yo desarrollé una técnica de secuenciación masiva para la obtención de perfiles locus-específicos metilación del DNA que permitió el estudio en masa de múltiples L1HS individualmente usando secuenciación Illumina (Sanchez-Luque et al. Mol Cell 2019), y recientemente hemos desarrollado una técnica con mayor cobertura basada en Oxford Nanopore Technology (ONT) (Ewing et al. Mol Cell 2020). Con esto encontramos unos L1HS concretos que están hipometilados consistentemente tanto en tejidos humanos adultos como en un modelo de células madre embrionarias humanas (hESC), lo que sugiere que escapan el silenciamiento establecido en el embrión temprano y permanecen activos en tejidos adultos. Más aún, algunos coincidían en presentar una mutación para el sitio de unión del factor de transcripción Yin Yang 1 (YY1) típicamente asociado a pluripotencia. Yo postulo que YY1 o el sitio de unión a YY1 están involucrados en la ruta de señalización que identifica y silencia los L1HS durante la reprogramación embrionaria. De hecho, YY1 parece estar igualmente involucrado en el silenciamiento y/o metilación del DNA de otros retrotransposones y también retrovirus, lo que sugiere que existe un sistema de control central que es utilizado por elementos genéticos móviles tanto endógenos como foráneos (Bernhard et al. PLoS ONE 2013; Schlesinger et al. Cell Rep 2013).
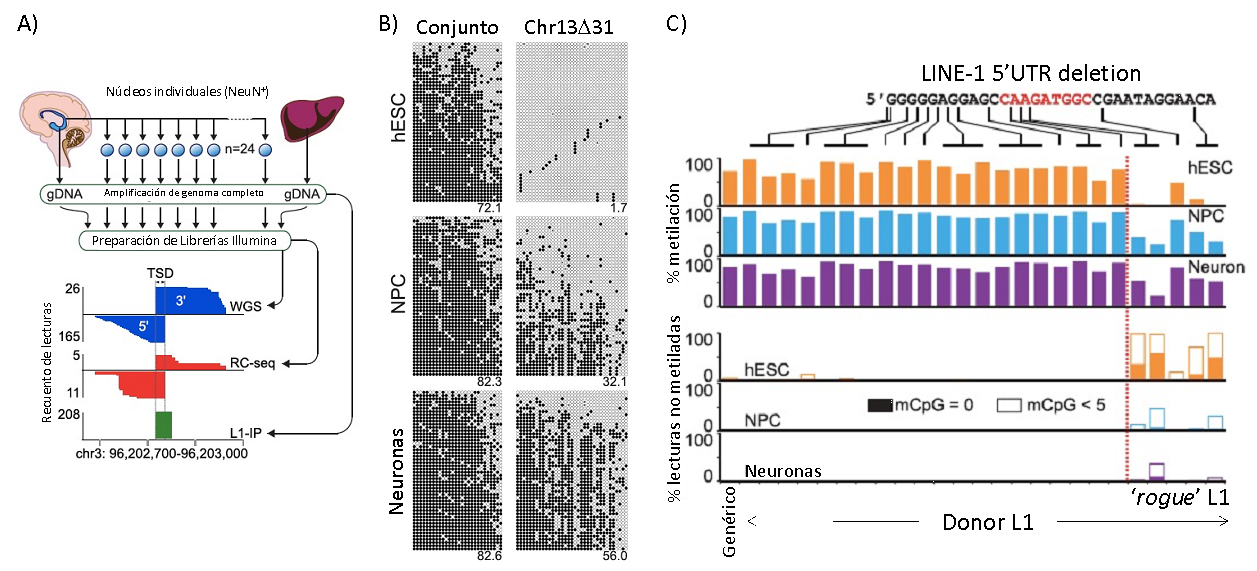
Figura 2. Retrotransposición de L1 y perfil epigenético en células madre embrionarias humanas (hESCs) y células diferenciadas (Adaptado de Sanchez-Luque et al. Mol. Cell 2019). A) Estrategia para la identificación de inserciones somáticas de L1 en neuronas de muestras humanas post-mortem mediante amplificación de genoma complete de single-neurons y una combinación de diferentes técnicas de secuenciación masiva (secuenciación de genoma completo directa, WGS; retrotransposon capture-sequencing, RC-seq; y L1-insertion profiling, L1-IP). B) Estudio de metilación del DNA de la población de L1 de forma genética y del locus específico Chr13D31 (donde se identificó un L1 'rogue') en hESCs y en progenitores neuronales (NPCs) y neuronas obtenidas por diferenciación in vitro. Cada panel contiene 50 lecturas (horizontal) donde los círculos negros y blancos representan CpG metiladas y no metiladas respectivamente. C) Estudio extensivo de la metilación del DNA en hESCs, NPCs y neuronas, de loci individuales de L1 con pequeños truncamientos en su 5'UTR. Los elementos con truncamientos de >14nt permanecen ampliamente hipometilados tras la diferenciación. En el panel inferior, la parte rellena y la parte vacía de las barras representan la fracción de lectura con ninguna o con <5 CpG metiladas, respectivamente.
En mi laboratorio, yo pretendo de caracterizar esta ruta de señalización involucrada en salvaguardar la integridad del genoma de la amenaza de los elementos genéticos móviles, al tiempo que identificar el DNA propio del extraño dentro de nuestro genoma y silenciar las integraciones de elementos foráneos como provirus. Para ello, tengo la intención de desarrollar una aproximación basada en edición del genoma por CRISPR-Cas9 de forma locus-específica para modificar loci de L1HS y testar el rol que tienen los distintos motivos de secuencia del su promotor en el silenciamiento epigenético. Esto se combinará con técnicas punteras de secuenciación (Illumina, ONT) y de biología molecular, y con el uso de modelos de cultivo de células madre embrionarias y retrovirus.
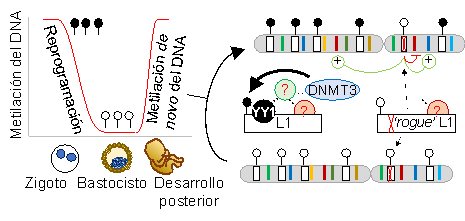
Figura 3. Hipótesis de investigación. La reprogramación celular durante el desarrollo embrionario temprano implica la desmetilación general del DNA, seguida de una remetilación de los elementos L1 alrededor del estadio de blastocisto (círculos negros y blancos representan CpGs metiladas y no metiladas respectivamente). YY1 y otros factores candidatos estarían involucrados en reconocer secuencias dentro del promotor de L1 y reclutar la maquinaria de silenciamiento, que en último lugar conlleva la metilación de novo del DNA. Copias de L1 mutantes que no son reconocidas por dichos factores, permanecerían hipometiladas (?rogue?) y se movilizarían en células humanas.
PUBLICATIONS THAT SUPPORT THIS RESEARCH LINE
1 Smits N, Rasmussen J, Bodea GO, Amarilla AA, Gerdes P, Sanchez-Luque FJ, Ajjikuttira P, Modhiran N, Liang B, Faivre J, Deveson IW, Khromykh AA, Watterson D, Ewing AD, Faulkner GJCA 2021. No evidence of human genome integration of SARS-CoV-2 found by long read DNA sequencing. Cell Reports 7;109530 [Q1; IF 9.423; 3 citations].
2 Ewing ADCA, Smits N, Sanchez-Luque FJ, Faivre J, Brennan PM, Richardson SR, Cheetham SW, Faulkner GJCA 2020. Nanopore sequencing enables comprehensive transposable element epigenomic profiling. Molecular Cell 80;915-28 [D1; IF 17.970; 33 citations].
3 Microbiology Resource Announcements 9;e00453-20 [cit. 2].
4 Sanchez-Luque FJCA, Kempen MHC, Gerdes P, Vargas-Landin DB, Richardson SR, Troskie R-L, Jeasuadian JS, Cheetham SW, Carreira PE, Salvador-Palomeque C, Garcia-Canadas M, Munoz-Lopez M, Sanchez L, Lundberg M, Macia A, Heras SR, Brennan PM, Lister R, Garcia-Perez JL, Ewing AD, Faulkner GJ.CA 2019. LINE-1 Evasion of Epigenetic Repression in Humans. Molecular Cell 75;590-604 [D1; IF 17.970, 52 citations].
5 Salvador-Palomeque C*, Sanchez-Luque FJ*, Fortuna PRJ, Ewing AD, Wolvetang EJ, Richardson SRCA, Faulkner GJCA. 2019 Dynamic Methylation of an L1 Transduction Family during Reprogramming and Neurodifferentiation. Molecular and Cellular Biology 39;e00499-18 [Q2; IF 4.272; 8 citations]. *co-first author.
6 Nguyen THM*, Carreira PE*, Sanchez-Luque FJ*, Schauer SN, Fagg AC, Richardson SR, Davies CM, Jesuadian JS, Kempen M-JHC, Troskie R-L, James C, Beaver EA, Wallis TP, Coward JIG, Chetty NP, Crandon AJ, Venter DJ, Armes JE, Perrin LC, Hooper JD, Ewing AD, Upton KRCA, Faulkner GJCA 2018. L1 Retrotransposition Heterogeneity in Ovarian Tumor Cell Evolution. Cell Reports 23;3730-40 [Q1; IF 9.423; 33 citations]. *co-first author.
7 Schauer SN, Carreira PE, Shukla R, Gerhardt DJ, Gerdes P, Sanchez-Luque FJ, Nicoli P, Ghisletti S, Dos Santos A, Rapound D, Samuel D, Faivre J, Ewing ADCA, Richardson SRCA, Faulkner GJCA 2018. L1 retrotransposition is a common feature of mammalian hepatocarcinogenesis. Genome Research 28;639-53 [D1; IF 9.043; 52 citations].
8 Richardson SRCA, Gerdes P, Gerhardt DJ, Sanchez-Luque FJ, Bodea G-O, Munoz-Lopez M, Jesuadian JS, Kempen M-JHC, Carreira PE, Jeddeloh JA, Garcia-Perez JL, Kazazian HH Jr, Ewing AD, Faulkner GJCA 2017. Heritable L1 retrotransposition in the mouse primordial germline and early embryo. Genome Research 27;1395-405 [D1; IF 9.043, 70 citations].
9 Upton KU, Gerhardt DJ, Jesuadian JS, Richardson SR, Sanchez-Luque FJ, Bodea G-O, Ewing AD, Salvador-Palomeque C, van der Knaap MS, Brennan PM, Vanderver A, Faulkner GJCA 2015. Ubiquitous L1 mosaicism in hippocampal neurons. Cell 161;228-39 [D1; IF 41.584; 280 citations].
ORGANISMOS FINANCIADORES ÚLTIMOS 5 AÑOS
|