|
SUMMARY OF RESEARCH
RESEARCH BACKGROUND - MOBILE DNA IN HUMANS
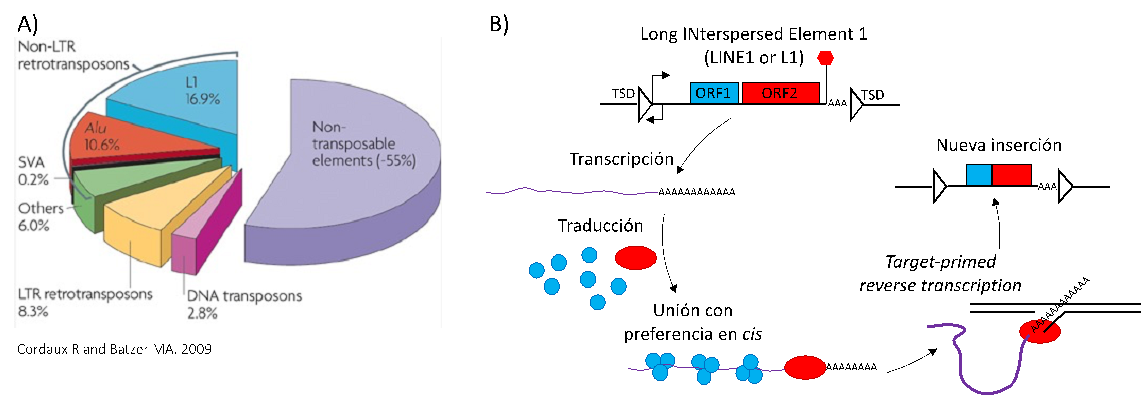
The human genome has been subject of many waves of colonisation by mobile DNA elements that have successively multiplied and declined along evolution, leaving uncountable fossil copies as consequence of their past replicative activity. These copies represent about half of the genome size and are now consolidated and repurposed as structural, regulatory and even occasionally coding sequences (Lander et al. Nature 2001). Within this polyphyletic group of elements, only a group of retrotransposons are the latest active in humans. They are transcribed into an intermediary RNA that is reverse transcribed back to DNA into a new copy (Kazazian. FT Press Science 2011). In this group, endogenous retroviruses (ERV; closely related to retroviruses) were active in humans until very recently (and still active in other organisms including drosophila, mouse and pigs) (Belshaw et al. J Virol 2005; Maksakova et al. PLoS Genet 2006). Only the non-ERV elements LINE-1 (L1), Alu and SVA are currently mobile in humans, causing insertional mutagenesis in both the germline and somatic cells, although limited by epigenetic restriction. They create human genetic diversity at a rate of ~1 heritable insertion in every 40 births, occasionally leading to cases of genetic disease perhaps within a tolerable evolutionary scale (Ewing and Kazazian. Genome Res 2010; Feusier et al. Genome Res 2019; Kazazian and Moran. N Engl J Med 2017). However, we and others have shown they get dysregulated in cancer, where their input in genetic diversity influences tumour progression and metastasis (Scott et al. Genome Res 2016; Tubio et al. Science 2014; Nguyen et al. Cell Rep 2018).
A functional human L1 copy is a ~6kb sequence that encodes instrumental activities for its mobilisation, including AP endonuclease (EN) and reverse transcriptase (RT). This mechanism alternates DNA and RNA vessels and is probably rooted in the evolutionary origins of the current nucleic acid?s information flow (Richardson et al. Mob DNA III 2015). The 5?UTR with internal Pol_II promoter activity drives sense transcription of a L1 polyadenylated mRNA, which is translated in the cytoplasm and bound in cis by the mobilisation machinery. They return to the nucleus to synthesise the new copy by target-primed reverse transcription (TPRT), which involves a single strand nick in the genomic insertion site and the use of the nick 3?hydroxyl end as primer for reverse transcribing L1 mRNA into the first cDNA strand (Luan et al. Cell 1993; Cost et al. EMBO J 2002). It is unclear how L1 restores the genome integrity of this strand and synthesises the complementary cDNA, but probably involves the coordination of cellular machinery. Conversely, Alu and SVA are recently evolved, smaller and non-coding composite elements that mobilise by hijacking L1 machinery in trans (Dewannieux et al. Nat Genet 2003; Hancks et al. Hum Mol Genet 2011).
Figure 1. Mobile Genetic Elements abundance in humans and mobilisation mechanism. A) Pie chart with the abundance of the major families of mobile genetic elements or transposable elements in the human genome (Cordaux and Batzter 2009). B) Mobilisation mechanism of L1 elements, which involves full-length L1 translation into a mRNA that is translated in the cytoplasm; binds in cis to its own mobilisation machinery; and returns to the nucleus, where the ORF2p endonuclease and reverse transcriptase catalyses the synthesis of a new copy by target-primed reverse transcription.
EPIGENETIC CONTROL OF FOREIGN AND MOBILE DNA IN THE HUMAN GENOME
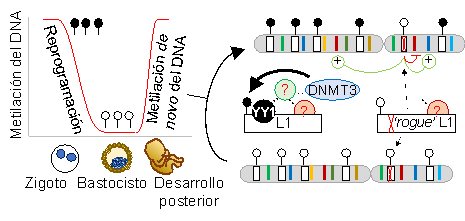
Humans and L1 have co-evolved to achieve a mobilisation rate compatible with their common survival. Indeed, there are several past and present examples of mobile DNA positively influencing their hosts, even in humans (Kapitonov et al. PLoS Biol 2005). A major mechanism controlling the activity of L1HS, the youngest and only mobilisation-competent L1 family in humans, is the epigenetic silencing of their promoter, which is mediated by DNA methylation of a CpG island present in the 5?UTR (Muotri et al. Nature 2010). However, genomewide DNA de-methylation occurs in the early embryo and the primordial germline in two separate waves of reprogramming (Greenberg and Bourc?his. Nature 2019). This is apparently needed for embryonic cells to achieve pluripotency at the early stages of development, but also enables a window of L1 activation that allow it to reach the germline genome and perpetuate new copies. Notably, there are >330 different full-length L1HS genomic copies in every human, each with a promoter with transcriptional potential. Approximately 10% of the full-length L1HS are polymorphic, non-fixed in the population, and add to the genetic diversity in humans (Ewing and Kazazian. Genome Res. 2011). Most L1HS are quickly silenced by de novo DNA methylation before the pre-implantation of the embryo, although the mechanism by which the cells localise and silence during reprogramming remains unknown.
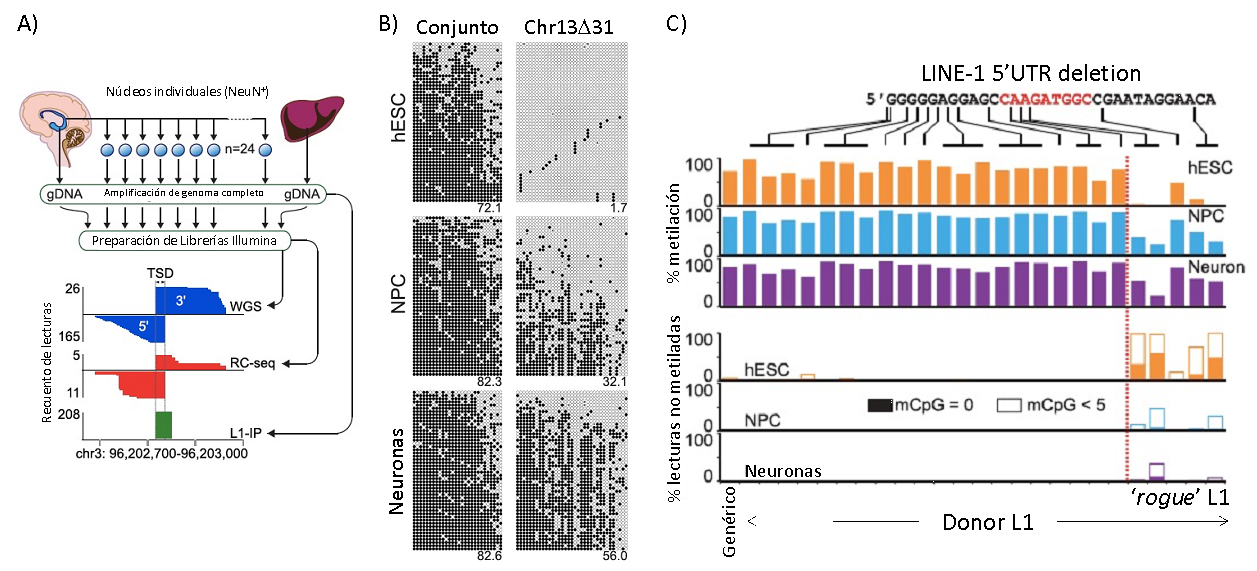
L1HS copies have been typically studied in bulk due to their extremely high sequence identity. Their recent and replicative origin has traditionally hindered their individual loci with omic techniques based in short-read sequencing. During my postdoct at the Faulkner lab (Mater Research Institute, Brisbane, Australia), I developed a high-throughput technique for locus-specific DNA methylation profiling that allowed the en masse study of L1HS individually using Illumina sequencing (Sanchez-Luque et al. Mol Cell 2019), and recently we have developed Oxford Nanopore Technology (ONT)-based approach with higher coverage (Ewing et al. Mol Cell 2020). We found that specific L1HS are recurrently hypomethylated in human adult tissues as well as in a model of human embryonic stem cells (hESCs), suggesting that these elements escape early embryonic silencing and remain active in adult tissues. We noticed that a subset of them share a specific deletion affecting a binding site for the transcription factor Yin Yang 1 (YY1), typically associated to pluripotency. I speculate that YY1 or the YY1-binding site are involved in the signalling pathway identifying and silencing L1HS during reprogramming. Notably, YY1 appears to be involved in silencing and/or DNA methylation of other retrotransposons and also retroviruses, suggesting a core regulatory pathway used by endogenous and foreign mobile elements (Bernhard et al. PLoS ONE 2013; Schlesinger et al. Cell Rep 2013).
Figure 2. L1 retrotransposition and epigenetics in human embryonic stem cells (hESCs) and differentiated cells (Adapted from Sanchez-Luque et al. Mol. Cell 2019). A) Strategy for identifying somatic L1 insertions in human post-mortem neurons by single-neurons whole genome amplification and a combination of high-throughput sequencing techniques (direct whole genome sequencing, WGS; retrotransposon capture-sequencing, RC-seq; L1-insertion profiling, L1-IP). B) DNA methylation profiling of the bulk LINE-1 population and the specific locus Chr13D31 (where a 'rogue' L1 was identified) in hESCs and in vitro differentiated neuronal progenitor cells (NPCs) and neurons. Each panel contains 50 random reads (horizontal) where black and white circles represent methylated and unmethylated CpGs (met-CpG) respectively. C) Summary of an extensive profiling of multiple individual locus-specific L1s in hESCs, NPCs and neurons with different small truncations in their 5'UTR. Elements truncated >14nt remain largely unmethylated upon differentiation. In the lower panel, solid and empty bar sections indicate reads (as in B) with no or <5 met-CpGs respectively.
In my lab, I intend to characterise this regulatory pathway involved in safeguarding genome integrity from endogenous mobile DNA threats, but also in identifying self and non-self DNA within our genome and silencing integrations of foreign elements including proviruses. I intend to develop a locus-specific CRISPR-Cas9 genome editing approach and modify specific L1HS loci in order to challenge the role in silencing of sequence motifs in their promoter. This will be combined with state-of-the-art sequencing (Illumina, ONT), molecular biology technologies, the use of human embryonic stem cell culture and retrovirus culture models.
Figure 3. Research hypothesis. Early embryonic reprogramming involves general DNA demethylation followed by remethylation of L1 elements around blastocyst stage (black and white lollipops represent methylated and unmethylated CpGs). YY1 and other candidate factors may be involved in binding L1 promoter sequence and recruit silencing machinery, which ultimately mediates de novo DNA methylation. Mutant copies with compromised binding for these factors may remain hypomethylated (rogue) and retrotranspose in human cells.
PUBLICATIONS THAT SUPPORT THIS RESEARCH LINE
1 Smits N, Rasmussen J, Bodea GO, Amarilla AA, Gerdes P, Sanchez-Luque FJ, Ajjikuttira P, Modhiran N, Liang B, Faivre J, Deveson IW, Khromykh AA, Watterson D, Ewing AD, Faulkner GJCA 2021. No evidence of human genome integration of SARS-CoV-2 found by long read DNA sequencing. Cell Reports 7;109530 [Q1; IF 9.423; 3 citations].
2 Ewing ADCA, Smits N, Sanchez-Luque FJ, Faivre J, Brennan PM, Richardson SR, Cheetham SW, Faulkner GJCA 2020. Nanopore sequencing enables comprehensive transposable element epigenomic profiling. Molecular Cell 80;915-28 [D1; IF 17.970; 33 citations].
3 Microbiology Resource Announcements 9;e00453-20 [cit. 2].
4 Sanchez-Luque FJCA, Kempen MHC, Gerdes P, Vargas-Landin DB, Richardson SR, Troskie R-L, Jeasuadian JS, Cheetham SW, Carreira PE, Salvador-Palomeque C, Garcia-Canadas M, Munoz-Lopez M, Sanchez L, Lundberg M, Macia A, Heras SR, Brennan PM, Lister R, Garcia-Perez JL, Ewing AD, Faulkner GJ.CA 2019. LINE-1 Evasion of Epigenetic Repression in Humans. Molecular Cell 75;590-604 [D1; IF 17.970, 52 citations].
5 Salvador-Palomeque C*, Sanchez-Luque FJ*, Fortuna PRJ, Ewing AD, Wolvetang EJ, Richardson SRCA, Faulkner GJCA. 2019 Dynamic Methylation of an L1 Transduction Family during Reprogramming and Neurodifferentiation. Molecular and Cellular Biology 39;e00499-18 [Q2; IF 4.272; 8 citations]. *co-first author.
6 Nguyen THM*, Carreira PE*, Sanchez-Luque FJ*, Schauer SN, Fagg AC, Richardson SR, Davies CM, Jesuadian JS, Kempen M-JHC, Troskie R-L, James C, Beaver EA, Wallis TP, Coward JIG, Chetty NP, Crandon AJ, Venter DJ, Armes JE, Perrin LC, Hooper JD, Ewing AD, Upton KRCA, Faulkner GJCA 2018. L1 Retrotransposition Heterogeneity in Ovarian Tumor Cell Evolution. Cell Reports 23;3730-40 [Q1; IF 9.423; 33 citations]. *co-first author.
7 Schauer SN, Carreira PE, Shukla R, Gerhardt DJ, Gerdes P, Sanchez-Luque FJ, Nicoli P, Ghisletti S, Dos Santos A, Rapound D, Samuel D, Faivre J, Ewing ADCA, Richardson SRCA, Faulkner GJCA 2018. L1 retrotransposition is a common feature of mammalian hepatocarcinogenesis. Genome Research 28;639-53 [D1; IF 9.043; 52 citations].
8 Richardson SRCA, Gerdes P, Gerhardt DJ, Sanchez-Luque FJ, Bodea G-O, Munoz-Lopez M, Jesuadian JS, Kempen M-JHC, Carreira PE, Jeddeloh JA, Garcia-Perez JL, Kazazian HH Jr, Ewing AD, Faulkner GJCA 2017. Heritable L1 retrotransposition in the mouse primordial germline and early embryo. Genome Research 27;1395-405 [D1; IF 9.043, 70 citations].
9 Upton KU, Gerhardt DJ, Jesuadian JS, Richardson SR, Sanchez-Luque FJ, Bodea G-O, Ewing AD, Salvador-Palomeque C, van der Knaap MS, Brennan PM, Vanderver A, Faulkner GJCA 2015. Ubiquitous L1 mosaicism in hippocampal neurons. Cell 161;228-39 [D1; IF 41.584; 280 citations].
FUNDING AGENCIES LAST 5 YEARS
|